") Italiano (IT)
Italiano (IT) ") English (UK)
English (UK) La genetica dei mantelli
Abbiamo parlato in un altro articolo dei possibili aspetti del mantello del gatto, con particolare attenzione al Maine Coon. Tutta questa varietà di colori, in realtà, è governata da poche semplici leggi. Bisogna però conoscere alcune nozioni di base della genetica per comprendere come funzionano. Il discorso potrà sembrare un pochino complicato, ma non scoraggiatevi!
 Le caratteristiche di ogni essere vivente sono stabilite dal corredo genetico, a cui contribuiscono in ugual misura il padre e la madre. Il corredo genetico è costituito da coppie di cromosomi identici, schematizzabili come due X, tranne per quello che determina il sesso. Infatti, in questo caso, uno dei due cromosomi perde una "zampetta", e passa dalla forma a X a una forma a Y. Quando viene generato un nuovo essere vivente, il suo sesso è femminile se la combinazione è XX, maschio se la combinazione è XY.
Le caratteristiche di ogni essere vivente sono stabilite dal corredo genetico, a cui contribuiscono in ugual misura il padre e la madre. Il corredo genetico è costituito da coppie di cromosomi identici, schematizzabili come due X, tranne per quello che determina il sesso. Infatti, in questo caso, uno dei due cromosomi perde una "zampetta", e passa dalla forma a X a una forma a Y. Quando viene generato un nuovo essere vivente, il suo sesso è femminile se la combinazione è XX, maschio se la combinazione è XY.
 Il modo con cui si combinano i geni si visualizza spesso con il cosiddetto quadrato di Punnet. Visto che è uno strumento semplice ma essenziale per lavorare con la genetica, vediamo come si costruisce nel caso della determinazione del sesso. Basta disegnare una griglia 2x2, e sul lato superiore scrivere i geni della madre, e su quello sinistro quelli del padre. Ovviamente, si può fare anche viceversa. Quindi in ogni quadrato di ogni colonna della griglia si riporta il gene corrispondente a quella colonna, in questo caso la X rosa della madre in prima colonna e di nuovo la X rosa della madre in seconda. Poi, in ogni quadrato della riga, si trascrive il gene relativo a quella riga, cioè la X blu del padre e la Y blu del padre. Nei singoli quadrati della griglia ottienamo quindi le possibili combinazione dei geni di padre e madre. La combinazione XX indica una femmina, quella XY indica un maschio. In questo esempio, risulta chiaro che la probabilità di avere un figlio maschio o femmina è del 50%, ma va assolutamente sottolineato che questa probabilità comincia ad aver senso solo per un elevato numero di eventi, cioè di cuccioli nella stessa cucciolata, all'incirca a partire da 10. Poiché ogni evento (nascita) è indipendente da ogni altro, la statistica totale comincia ad aver senso solo per numeri molto elevati di parti, diciamo, per fissare le idee, una decina. Questa considerazione è valida per ogni valutazione statistica, ed è importante tenerlo bene a mente.
Il modo con cui si combinano i geni si visualizza spesso con il cosiddetto quadrato di Punnet. Visto che è uno strumento semplice ma essenziale per lavorare con la genetica, vediamo come si costruisce nel caso della determinazione del sesso. Basta disegnare una griglia 2x2, e sul lato superiore scrivere i geni della madre, e su quello sinistro quelli del padre. Ovviamente, si può fare anche viceversa. Quindi in ogni quadrato di ogni colonna della griglia si riporta il gene corrispondente a quella colonna, in questo caso la X rosa della madre in prima colonna e di nuovo la X rosa della madre in seconda. Poi, in ogni quadrato della riga, si trascrive il gene relativo a quella riga, cioè la X blu del padre e la Y blu del padre. Nei singoli quadrati della griglia ottienamo quindi le possibili combinazione dei geni di padre e madre. La combinazione XX indica una femmina, quella XY indica un maschio. In questo esempio, risulta chiaro che la probabilità di avere un figlio maschio o femmina è del 50%, ma va assolutamente sottolineato che questa probabilità comincia ad aver senso solo per un elevato numero di eventi, cioè di cuccioli nella stessa cucciolata, all'incirca a partire da 10. Poiché ogni evento (nascita) è indipendente da ogni altro, la statistica totale comincia ad aver senso solo per numeri molto elevati di parti, diciamo, per fissare le idee, una decina. Questa considerazione è valida per ogni valutazione statistica, ed è importante tenerlo bene a mente.
Su ogni coppia di cromosomi, ci sono sequenze di decine di migliaia di molecole che determinano tutte le caratteristiche dell'essere vivente. Ogni sequenza del cromosoma controlla una singola funzione, per esempio una sequenza la forma degli occhi, un'altra la lunghezza del femore, ecc. ecc. Queste singole sequenze prendono il nome di geni.
Le coppie di cromosomi sono uguali funzionalmente, nel senso che ogni cromosoma della coppia ha la stessa sequenza di geni, e ognuno dei due geni sui due cromosomi della coppia controlla esattamente la stessa funzione. I geni però non sono esattamente identici, poiché uno dei cromosomi della coppia deriva dalla madre, e uno dal padre, e possono quindi differire per il loro stato. I geni infatti possono essere in due stati, che per semplicità chiameremo "acceso" o "spento". Se il gene è acceso, e questo si esprime indicando il gene con la lettera maiuscola, la funzione corrispondente è attiva, se è spento, la funzione corrispondente non è attiva.
Dalla combinazione dei due geni nella coppia di cromosomi, risulta l'azione complessiva della funzione. Per un gene generico G ci possono quindi essere le combinazione GG (entrambi geni accesi) Gg o gG (uno acceso, uno no), e quella gg (entrambi i geni spenti). Il gene acceso viene indicato come gene dominante, quello spento come recessivo. Per esprimere una caratteristica legata a un gene dominante, basta che sia presente il gene corrispondente. In altre parole, se il nostro gene ipotetico controlla la presenza degli occhi verdi, tutti i gatti nel cui corredo genetico sia presente la combinazione GG, Gg o gG avranno gli occhi verdi. La combinazione GG è detta omozigote, poiché i geni sono nello steso stato, quella Gg o gG eterozigote, poiché i geni sono in due stati diversi. Le due combinazioni Gg e gG sono esattamente equivalenti e quindi se ne indica solo una. La combinazione gg invece, anch'essa omozigote, da luogo a un colore non-verde. Questa affermazione un po' strana dice solo che il colore degli occhi del gatto dipenderà dal resto del patrimonio genetico, ma che di sicuro non sono verdi.
Quindi, per ricapitolare, una particolare caratteristica legata a un gene dominante si manifesterà nel 75% dei casi (sempre assumendo un numero di eventi molto elevato, al limite infinito), quella legata al gene recessivo nel 25%. Nel caso del gene dominante però, solo un soggetto su tre sarà omozigote, gli altri due si porteranno nel loro patrimonio genetico il gene recessivo, che si potrà manifestare nelle future generazioni se si combina con un altro gene recessivo. Diventa importante quindi distinguere tra genotipo, cioè l'insieme del corredo genetico, e fenotipo, cioè l'insieme delle caratteristiche esterne che si manifestano.
Chiarito il ruolo dei geni, possiamo adesso vedere come questo influenza il colore del mantello dei gatti. Il mantello originario del gatto è nero tigrato mackerel, quindi con ogni pelo a bande alternate chiare e scure (agouti) e tigratura a linee sottili verticali, dove si alternano zone con pelo a bande e zone con pelo uniforme. Il colore nero si esprime grazie a un gene Black, dominante. Nei Maine Coon, l'unica combinazione ammessa è quella eterozigote dominante BB. Nelle altre razze, come ad esempio il Sacro di Birmania, sono presenti anche altre varianti, legate al fatto che B ha due geni recessivi possibili, b e bl. Il primo, nella combinazione omozigote recessiva bb, da il colore chocolate, il secondo blbl da il colore cinnamon (cannella).
Nel corso dell'evoluzione, una mutazione genetica ha fatto sì che il pigmento nero tipico del gatto ancestrale si tramutasse, sotto l'azione di nuovo gene, in un pigmento rosso. Questo effetto, gestito dal gene mutato Orange, rende rosse le zone del pelo che in origine erano nere. In altre parole, nel pelo non c'è più un'alternanza di banda nere e chiare, ma di bande rosse e chiare. Questo gene, quando è acceso (quindi O grande), copre l'azione del gene Black, quindi è in grado di trasformare il nero in rosso anche nella sua espressione eterozigote Oo (O grande o piccolo). Questa azione di copertura tecnicamente si chiama epistasi (dal greco epistasis, posizione dominante), poiché un gene maschera l'azione di un'altro gene. Nella sua espressione omozigote oo (o piccolo o piccolo), il gatto rimane non-rosso, e quindi nero. La Natura però ha voluto farci un bello scherzo, e ha localizzato questo nuovo gene mutato Orange proprio sulla zampa del cromosoma sessuale X che viene persa quando si trasforma in Y. Quindi, il gene per il rosso è portato solo dal cromosoma X, ed è allora un gene legato al sesso. In questo caso, il quadrato di Punnet si complica un pochino, poiché va tenuto presente anche il sesso del gatto. Vediamo cosa succede mettendo insieme un maschio nero XY o- e una femmina rossa XX OO.
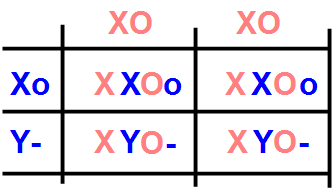
Innanzitutto, vediamo che il gatto nero ha un solo gene per il colore rosso, nella sua forma recessiva o. L'altro gene della coppia è mancante, poiché il maschio ha un solo cromosoma X, e si indica con un trattino -. La femmina invece contribuisce con entrambi i cromosomi O. Guardiamo innanzitutto la seconda riga del quadrato, dove c'è solo la combinazione dei cromosomi sessuali XY. I maschi che nascono da questo accoppiamento sono entrambi rossi (O-), quindi ereditano il colore della madre. Sulla prima riga ci sono invece solo le combinazioni XX, quindi femmine, che hanno entrambi i geni O e o, uno ereditato dalla madre, uno dal padre. DI che colore saranno queste femmine Oo? In teoria, la O è dominante, quindi dovrebbero essere rosse. In pratica, avviene un fenomeno particolare durante la fase di crescita dell'embrione, legato al fatto che il gene Orange è sul cromosoma sessuale X. Infatti, mentre le femmine hanno due cromosomi X, il maschio ne ha uno solo, e quindi tutti gli altri geni presenti sul cromosoma X hanno, nel maschio, una possibilità di espressione genica dimezzata. In parole semplici, i geni del maschio sul cromosoma X produrrebbero la metà delle proteine dei geni della femmina, con il risultato di un svantaggio competivo evidente. Madre Natura ha posto rimedio a questo problema facendo sì che, in tutte le cellule della femmina, durante lo sviluppo embrionale uno dei due cromosomi X si disattivi, in maniera assolutamente casuale cellula per cellula e, per tutte le cellule, in un momento casuale durante lo sviluppo del feto. Il risultato, dal punto di vista del mantello, è che le cellule responsabili della produzione del pigmento del pelo potranno produrre o pigmento rosso, o pigmento nero, dando luogo a quella mescolanza di colori rosso e nero tipico delle tortie. Se l'inattivazione avviene precocemente, quando ancora le cellule sono poche, le duplicazioni successive di una cellula in cui è stato attivato uno dei due geni, diciamo quello per il colore rosso, mantengono lo stesso colore, e analogamente per il nero. Questo produce un mantello a larghe macchie rosse e nere. Se invece l'inattivazione avviene tardi, i due colori appaiono ben mescolati. Quindi, ricapitolando, dall'accoppiamento tra un maschio nero e una femmina rossa, nascono femmine tortie e maschi rossi.
L'espressione dei colori nero e rosso attraverso i geni Black e Orange può essere modificata dall'azione del gene Dense. Nella condizione più comune, con la coppia DD, il gatto ha colore nero o rosso. Nella sua versione omozigote recessiva dd, la distribuzione del pigmento nei peli diventa disomogenea, con il risultato di avere maggior riflessione della luce e quindi un colore più diluito, meno intenso. Il corredo genetico BB-oo-dd dà luogo a gatti color grigio-blu, mentre quello BB-OO-dd a gatti color crema, e quello BB-Oo-dd a tortie blue-crema.
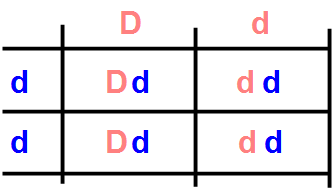
Il fatto che il colore diluito si manifesti solo quando il gene Dense è nella sua forma omozigote recessiva dd fa sì che sia necessario accoppiare gatti entrambi con colore diluito per ottenere cuccioli con colore diluito. Questo può però avvenire anche se uno dei genitore non esprime il gene della diluzione, ma lo porta nel suo patrimonio genetico, frutto per esempio di un ascendente. Infatti, accoppiando un Dd con un dd, dal quadrato di Punnet relativo si vede che, statisticamente, la metà dei cuccioli ha colore diluito, e la metà no, anche se porta con sé il gene recessivo relativo.
Un altro gene epistatico, cioè che maschera l'azione di altri geni, è quello legato alla presenza di macchie bianche nel mantello. Questo gene, detto piebald Spotting (a macchie pezzate), maschera in quelle zone l'espressione dei geni Orange e Black. Questo gene agisce durante lo sviluppo embrionale, impedendo alle cellule che generano il pigmento di raggiungere la loro posizione. Se sulle aree dove il gene agisce è presente del pelo, questo pelo risulta bianco. Se invece c'è pelle, come sul naso o sui polpastrelli, questa viene depigmentata a rosa. Il gene è dominante, anche se la sua espressione è variabile. Ci sono forti indicazioni che la versione dominante omozigote SS dia aree bianche di maggiore estensione rispetto alla versione eterozigote Ss, per quanto la quantità e la localizzazione delle macchie dei figli a partire da quella dei genitori sia al momento impredicibile. La versione omozigote recessiva ss ovviamente non dà macchie bianche.
Una versione rinforzata del gene S è quella del gene White, che da luogo a un mantello completamente bianco. Si tratta di un gene dominante, quindi sia la combinazione WW che quella Ww danno lo stesso risultato. Il meccanismo di azione è il medesimo del gene S, solo che interessa tutto il mantello del gatto. Si manifestano però degli altri effetti. Infatti, le cellule embrionali che originano il pigmento della pelle generano anche il pigmento del fondo dell'occhio e le cellule pigmentate dell'orecchio interno. Se l'azione del gene, nella fase della separazione iniziale delle cellule, si rivolge alle cellule responsabili della formazione del tessuto oculare, l'inibizione della pigmentazione si traduce nel fatto che quell'occhio avrà, al termine della crescita, colore blu o azzurro chiaro. Naturalmente, può succedere che un occhio sia depigmentato e l’altro no, dando luogo agli affascinanti occhi impari. Se l'azione del gene colpisce anche i tessuti auricolari, si può avere malfunzionamento o sordità, mono o bilaterale. Non ci sono comunque certezze in questo senso, e le regole di ereditarietà del colore degli occhi e della sordità sono ancora da stabilire, e solo qualche indicazione statistica ci può aiutare.
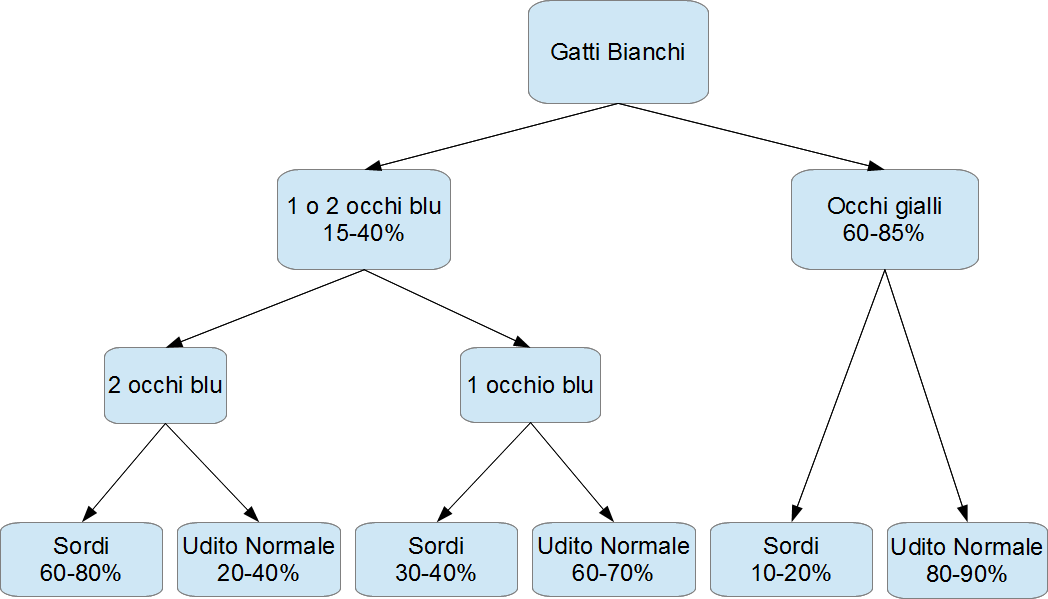
La prima cosa da notare è che se il cucciolo ha delle macchie colorate sul capo, una traccia del colore mascherato dal gene W, la probabilità che sia sordo è attorno al 30%, mentre sale al 50% se le macchie non ci sono. La possibile spiegazione è che se sono arrivate a destinazione un po' di cellule del pigmento del pelo, è plausibile che sia accaduto lo stesso per quelle dell'orecchio interno, sufficienti talvolta a evitare la sordità. La seconda correlazione è quella con il colore degli occhi.
Sulla base di una serie di studi condotti in varie nazioni, e visualizzati nella figura a sinistra, gatti con entrambi gli occhi gialli hanno una probabilità di sordità del 10-20%, gatti con occhi impari probabilità del 30-40%, gatti con entrambi gli occhi blu probabilità di sordità attorno al 60-80%. Si noti che quando si parla di sordità, essa può essere sia mono che bilaterale, equivalenti dal punto di vista genetico.
Finora ci siamo occupati implicitamente solo di gatti con colore solido, cioè senza nessuna tigratura. Come detto in un'altro articolo, il disegno della tigratura dipende da due condizioni; una è la presenza di pelo agouti, cioè in cui il singolo pelo non ha colore uniforme ma a bande alternate chiare e scure, l'altra è la presenza di bande alternate dove è presente pelo agouti (a bande alternate chiare e scure e quindi più chiaro) e pelo non-agouti (pelo unifome e quindi più scuro). Le zone dove il pelo è più scuro si chiamano markings (striature) della tigratura, Ciascuno di questi caratteri è controllato da un gene. II fatto che il pelo sia a bande oppure no dipende dal gene Agouti. Il gatto ancestrale ha il pelo agouti, e quindi l'espressione dominante AA è quella naturale, mentre la combinazione recessiva aa rende il pelo uniforme. Quindi nel gatto AA, oppure Aa (pelo agouti dominante) esiste il pelo a bande, e quindi sono possibili i disegni della tigratura, mentre il gatto aa non ha pelo agouti e non è (o non dovrebbe essere) tigrato. Il disegno della tigratura invece è gestito dal gene Tabby. Nella sua forma dominante omozigote TT da origine al disegno tabby mackerel, a righe parallele. La sua forma recessiva viene indicata con tb-tb, poiché luogo al disegno tabby classic, o blotched. La forma intermedia spotted non è dovuta a un gene particolare, ma a un insieme di più geni modificatori, non ancora identificati, che interrompono le linee parallele del mackerel e originano il disegno spotted. Gli stessi geni modificatori sono responsabili della dominanza incompleta del mackerel sul blotched. Il genotipo T-tb è spesso un mackerel non ben definito, con i due pattern che si mescolano tra loro. Per completezza, citiamo il gene Ticked Ta, che quando si esprime in forma dominante (TaTa o Tata) è epistatico (copre) tutti gli altri geni responsabili del disegno. Questo disegno della tigratura dà luogo per esempio al classico mantello del gatto Abissino.
Bisogna ricordare anche che il gene Agouti e il gene Tabby sono geni diversi, e che quindi un gatto solido porta con sé, a livello di genotipo, il gene della tigratura, qualunque sia, e che questo si può esprimere di nuovo qualora la progenie fosse tigrata. Inoltre, nei cuccioli solidi neri con gene Tabby non espresso, è comunque riconoscibile una tigratura fantasma, che spesso è ancora identificabile da adulto, guardando il pelo con luce radente. Nei gatti con gene Orange dominante poi, l'azione del gene non-agouti aa è assolutamente inefficace, tanto che è estremamente difficile avere gatti rossi solidi. La cosa è evidente anche nelle tortie tigrate, che hanno la tigratura molto più netta ed evidente nelle zone nere che in quelle rosse.
Come ultimo argomento, affrontiamo i colori silver, dove è presente uno sbiancamento della base del pelo, che può andare da un terzo a due terzi della lunghezza del pelo stesso. Nel primo caso, nei gatti con colore solido, si parla di colore smoke, nel secondo di shaded. Nei gatti tigrati, l'effetto silver è di schiarire la base del pelo nel marking delle tigrature, e rendere le bande del pelo color argento nelle zone con pelo agouti tra i markings. Nei Maine Coon, la teoria a un solo gene rende conto di quasi tutti gli aspetti fenotipici del mantello e quindi ci appoggeremo a quella. Il gene responsabile viene chiamato gene Inibitore della melanina, e nella sua espressione dominante II o Ii genera lo sbiancamento del pelo alla base, mentre nella versione recessiva ii non da luogo ad alcun effetto. Sulla base di questo semplice modello, è possibile prevedere la presenza del silver nella progenie, ma non rendere conto di tutti i dettagli, come il contrasto e l'estensione dello sbiancamento. I fattori genetici coinvolti nella produzione del silver ideale, solido o no, sono complessi. Non solo ci sono molti geni che interagiscono, ma i geni a volte non si esprimono pienamente, o sono in conflitto uno con l'altro. Ad esempio, il gene inibitore della melanina talvolta fa un pessimo lavoro nel bloccare la produzione del pigmento, risultando in un sottopelo troppo grigio, oppure con tonalità giallastre. Allo stesso modo, non-agouti mal espressi o sovra-espressione del gene inibitore della melanina possono generare un gatto dal mantello smoke pallido e slavato. Poligeni vari (insiemi di geni correlati), fattori legati all'influenza ambientale sull'espressione dei geni (epigenesi), o geni modificatori non ancora identificati si ritiene che portino a diversi fenotipi di colorazione, alcuni ritenuti più desiderabili rispetto ad altri da parte degli allevatori.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}